Feeling: the modal principle
The nervous system operates in two environments, an inner and an external one. It must direct the organism to act in the external environment so as to meet demands set and sensed in the inner environment; this activity is mediated by the external senses and by the skeletal muscle system. The nervous system also acts on the inner environment, sometimes directly through immediate control over respiration, heart beat, etc. and sometimes indirectly through its control of the endocrine system. Control over behavioral mode is the nervous system's basic means of co-ordinating activity in inner and outer environments. The brain is part of the inner environment.
Definition. Modal choice feeds the results of calculation back into the biological realm, activating brain regions and selecting programs of operation which commit the organism to an interpretation of its world.The concept of modal control has been explicated by Kilmer, McCulloch & Blum (1969) in an account of the reticular formation. They argue that animals must always be in one of several mutually exclusive modes of behavior and that the reticular formation, with its extensive afferent and efferent connections to the rest of the nervous system, is the obvious structure for implementing that commitment. The reticular formation facilitates activity in those brain regions which are most important for the current mode (see Fig. 1), while the actual behaviour of the organism when it is in the mode will be regulated by other brain centres and systems.
Kilmer et al. list 15 different modes, including, for example, sleeping, eating, fighting, hunting and grooming (see also MacLean, 1978). We are not interested in attacking or defending this particular list; what is important is recognizing that there is some small finite list of behavioural modes.
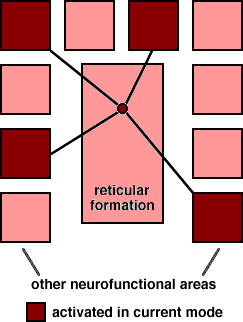
Fig. 1. Modal switching
This strategy for relating the modal principles to gross neuro-anatomy sees the brain as organized in layers about a central core (Pribram, 1971; 175 ff.). The central core, the reticular formation, sets the mode for the next layer, and each successive layer can be seen as setting the mode for the layers peripheral to it. Modes are effected by facilitating information processing in those areas of the brain which are most relevant to the corresponding behaviour and by turning control over to the appropriate neural centres (such as the limbic system and its evolutionary precursors; MacLean, 1978; Olds., 1977, Pribram, 1971). The selection of a particular behavioural goal, the formation of a plan to attain the goal and monitoring the success of that plan are all peripheral to the mode setting mechanism. Once the goal appropriate to a given mode has been attained, that mode will weigh less strongly in the reticular calculations which will then commit the brain to a different mode. The fundamental act is to commit the brain to a configuration of activity which interprets the external world as an arena for the satisfaction of particular inner needs, e.g. for nutrition, a sex partner, etc.
Biological principles are about maintaining the physical integrity of the organism, and, by implication, of the neural substrate which implements the informatic principles. Through the modal principle, the organism ultimately ensures that the schemas stored in other tissue have adaptive value (cf. Benzon, 1981). It does this through the implementation of the higher level modes of reorganization, play, imitation and speech (to be discussed in the section on 'Phylogeny'). We are therefore comfortable with Vanderwolf & Robinson's (1981) review of reticular function, which focuses on its role in regulating the activity of the cortex in a wide range of adaptive behaviours. The cortex implements the higher informatic functions and the reticular formation presides over their adaptive value.
The idea that differential activation of various brain areas subserves modal commitment is made particularly vivid by recent techniques for displaying patterns of blood flow in the cortex (Lassen, Ingvar & Skinhoj, 1978), which show differences in patterns of neocortical activation according to such behavioural modes as voluntary movement, speaking, reading silently, reading aloud. The linguistic modes are certainly unique to man. The important point here is that they are distinct behavioural modes, requiring different configurations of brain activity. Similar results have been obtained by measuring local metabolic rates in cortical areas mediating visual activity (Phelps, Kuhl & Mazziotta, 1981). It thus seems clear that the density of biochemical processes in a brain area varies according to demands being made on the area.
A rather different set of findings which can fruitfully be interpreted as clues to the biochemical implementation of the modal principle is the discovery of neuropeptides whose chemical structure is identical to those of gut hormones (Krieger & Liotta, 1979; Dismukes, 1979; Snyder, 1980; Bloom, 1981). What is particularly interesting is that these peptides have been discovered in brain regions which are apparently unrelated to the peripheral functions mediated by the corresponding gut hormone. Thus, for example, sex hormones have been found in the sonic motor nuclei of the oyster toadfish (Fine, Keefer & Leichnetz, 1982), and in auditory and vocal nuclei of South African clawed frogs (Kelley, 1980). In conjunction with evidence that neuronal local circuits are sensitive to chemical ambience (Schmitt, Dev & Smith, 1976), this suggests that modal control involves not only facilitating activity of various areas of the nervous system according to the demands of a given behavioural mode, but also a process of tagging or labeling stored patterns according to the modes which they serve (see Van Valen, 1982; see also Lewis et al ., 1981).
The modal function, the need to co-ordinate activity in the internal and external environments in commitment to a behavioural mode, thus can provide a way of interpreting two sets of phenomena: (1) gross patterns of activation in brain regions, and (2) the prevalence of neuropeptides which are identical to gut hormones and which might thus serve to label stored patterns. These two mechanisms can be seen as the means by which biological materials implement the psychological function of mode control.
Coherence: diagonalization
The technique of diagonalization was first employed by the mathematician Georg Cantor in his proofs that (1) the rational numbers are countable, and (2) the real numbers are not countable (Hermes & Markwald, 1974). The technique allowed Cantor to structure infinite sets in a way which made them tractable. The nervous system faces a similar problem. Sensory input is often ambiguous, with many interpretations possible and no obvious way of choosing among them. Hence the need for sensory coherence.
Definition. Diagonalization applies information from one channel to resolve ambiguity and impose structure in another channel.Perhaps the best way to grasp the principle is to consider a specific example first. We can then examine an explicit account of the principle. Gunnar Johansson's work on the perception of motion (1973, 1975) is suggestive. His basic technique has been to generate two-dimensional visual displays of dots in motion and ask his subjects what they see. One experiment involved images generated by moving people. Lights were attached to shoulders, elbows, wrists, hips, knees and ankles; and then films were made. These films show patterns of movement among 12 spots of light against a dark background. Subjects had little difficulty identifying the nature of the images, often doing so in a tenth of a second (the time required to project two frames of film). In fact, subjects performed better on this task than they did with mathematically simpler images derived from simple deformations and rotations of elementary geometric figures.
There is nothing surprising in the fact that we should be very sensitive to moving patterns generated by our fellow humans. Such sensitivity could be easily explained by evolutionary considerations. What is remarkable is the mechanism which realizes such sensitivity. There are infinitely many structures and motions which could generate the observed pattern of moving dots, yet the nervous system settles on one, the motion of a figure having members and joints arranged as humans do, and moving in the ways which humans do.
One way to interpret this phenomenon is to see it as an interaction between visual and kinesthetic space. Following the work of the Russian psychologist N. Bernstein (1967), Karl Pribram (1971) has argued that the motor cortex stores images of trajectories which it uses to regulate the activity of lower brain centres in generating movement. These images take the form of a Fourier transform of the trajectory (see also Gallistel, 1980). While Johansson uses vector geometry, it is clear that his results could be recast in Fourier terms, or that Bernstein's could be recast in vector geometrical terms. Hence, the same neural coding can be used for both the perception of organismic motion in the visual field and the generation of such motion in the kinesthetic field. The principle is that the analysis of information in one sensory channel must be consistent with the analysis of information in other sensory channels; and, perhaps more importantly, the analysis of information in one channel can only be made coherent through consideration of information in other channels. Diagonalization is a technique for achieving this coherence.
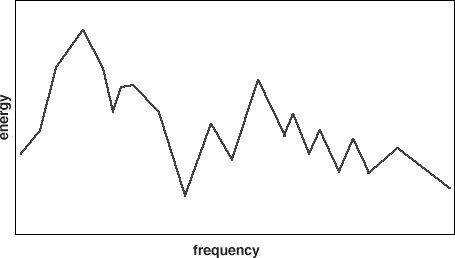
Fig. 2. A spectrum
To understand the application of diagonalization to this problem, we first introduce the concept of focal and residual analysis (a similar concept, attensity, can be found in Shaw & McIntrye, 1974). A perspicuous example comes from the history of astronomy (Kuhn, 1962; Hall, 1963). In the Ptolemaic scheme for planetary motion, the focus for analysing the path of a particular planet was a circle whose centre was the earth. The various cycles and epicycles which had to be added were the residuals needed to fully account for the observations. As the observations became more demanding, the residuals overwhelmed the focus. Consequently, Copernicus chose a new focus, a circle whose centre was the sun. This reduced the number of residuals needed by roughly a factor of two Kepler then introduced the ellipse as focal concept, achieving such precision that observed deviations could, in the context of Newtonian mechanics, be used to predict the existence of other planetary bodies.
The concept can be interpreted fairly naturally in the Fourier domain. The Fourier transform of a wave form yields a frequency spectrum, in which amplitude is graphed against frequency (see Fig. 2). The frequency with the highest amplitude is the focus of the transform, with the other frequency components being residual to it. However, if the signal is noisy and the sample finite, which is the situation the brain faces, there is more than one way to yield a plausible analysis. How does the system choose among them. If at least two channels of information about the stimulus are available, the system can diagonalize. The technique is simple (see Fig. 3): choose an analysis for each channel in which the components chosen to be focus and residuals, in order, exactly correspond in the two channels. This obviously requires that the coding in the two channels be compatible. In view of the evidence reviewed by Karl Pribram (1971, 1981; Pribram, Nuwer & Baron, 1974), some sort of coding in the frequency domain seems plausible, and we will assume it for the rest of this essay.
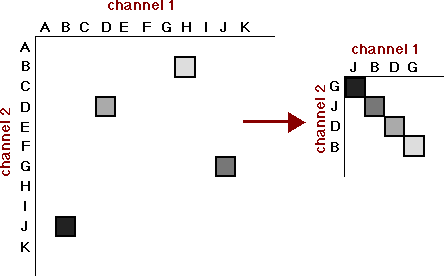
Fig. 3. Diagonalization
Now, let us consider some more examples. Piaget (1954) asserts that the object concept arises in infant experience through the co-ordination of information in various sensory channels. The senses do not display objects to the infant. Rather, objects are constructed by diagonalizing through the sensory information available. Edwin Land's (1977) retinex theory of colour vision is another example. Here, the system diagonalizes through the input in the three channels corresponding to the three types of cone receptor, with the attribution of colour to a region depending on that diagonalization. It seems to us that, in general, the perceptual constancies reflect various diagonalizing mechanisms.
We can also consider R. L. Gregory's (1966) report that people in cultures with visual environments which are very different from ours, such as the Zulu, perceive distances differently and are not subject to the same optical illusions. They are also unable to interpret drawings or photographs of familiar objects. Clearly, their visual systems have not been diagonalized for interpreting these types of visual stimuli (see the section 'Ontogeny'). Perhaps more dramatically, Gregory reports that people who are blind from birth but later recover vision, often through surgery, frequently require a long training period before they gain useful vision, and often enough their vision is never really useful. In one case, the patient's new visual capacities were best when directed toward objects of which he had previously had tactile experience. It would seem that the visual areas of the cortex matured without being diagonalized for visual stimuli. Hence, mere recovery of the peripheral channel was not sufficient to engender useful sight.
Now let us consider how diagonalization might be implemented in neural tissue; the work of Metzler & Spinelli (1977) on tilt constancy is suggestive. By recording activity in single cells in the visual cortex (area 17) of kittens, they established that some cells compensated for changes in head or body position while others did not. Metzler & Spinelli argued that the tilt constant cells are so because they are receiving information about head and body position from vestibular afferents. In our terms, the function of those cells within the visual system is to diagonalize through the input. Those cells provide the information needed to account for the structure of visual information to be attributed to the organism's own activity.
We would like to generalize from this to a consideration of Mountcastle's (1978) columnar hypothesis. The main idea is that the cortex is made up of a large number of processing columns which are perpendicular to the cortical sheet. The nature of the processing done in a given column depends on where its inputs are from--which subcortical or cortical regions. As one moves from primary, to secondary, to tertiary cortex (Luria's terms, 1973), the variety of columnar sensitivities, as defined by the range of inputs to the region, becomes more diverse (see also Lynch, 1980). The cortex is also organized into layers which are parallel to the cortical surface and hence orthogonal to the columns. Inputs from different broad classes of brain areas (e.g. the thalamus, basal ganglia, other cortical regions) terminate in different layers (Luria, 1973; Mountcastle, 1978; Szentagothai, 1978). The layers receiving subcortical input are relatively more developed in primary cortex, while those receiving cortical input are relatively more developed in the secondary and tertiary cortex. We thus suggest that the cortex is a diagonalizer, that each cortical region stores patterns which are diagonal functions of the inputs to the region.
Action: the decision principle
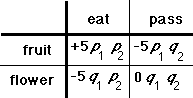
Fig. 4. A decision table
The decision principle is derived from the poetic principle of Roman Jakobson (1960), who was interested in the interplay between combinatory and selectional processes in the structure of poetic texts. But Jakobson's insight is more general. It can be seen as a particular type of minimax decision strategy (von Neumann & Morgenstern, 1944). Since it is generally impossible to maximize simultaneously on two variables, the idea is to choose so as to minimize the maximum likely loss. Consider the case of an animal trying to decide between eating and passing up the object before it (see Fig. 4). Eating is appropriate if the object is fruit, passing up if the object is a flower. Each cell contains the outcome value for the action along with the probabilities associated with identification of the object (p 1, q 1), which reflect the accuracy of the perceptual system, and with choice of an action (p 2, q 2), and which reflect current motivational state. Identifying the object as fruit or flower has no value to the animal independent of the probability of eating (hunger). Choosing to eat or pass up the object has no significance independent of the probability that the object is fruit (recognition). The loss intrinsic to each case (eating fruit, eating flower, etc.) has no significance independent of the probabilities. Action is unitary.
Definition. Decisions require the interaction of positive and negative feedback systems to minimax the dual ambiguity of perception and purpose.The organism sharpens the stimulus before it in a selectional array. It chooses which of the varieties of a given ontological type it has before it, thus determining its perceptual category--is the animal (type) a dog or cat (category)? And the organism establishes a sequence of actions which minimizes the difference between its current state of affairs and the desired goal (from the current mode) in a combinational array, thus establishing goals. Both of these processes can be constructed in closed loop systems. The primary evidence for a closed loop analysis of the nervous system is the ubiquity of reciprocal afferent and efferent connections between many classes of brain regions (Pribram, 1971; Eccles, 1973; Jones, 1974; Thatcher & John, 1977; Edelman & Mountcastle, 1978; Nauta & Feirtag, 1979).
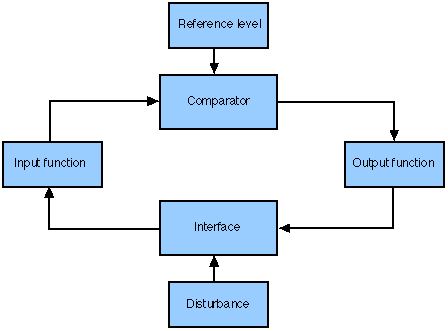
Fig. 5. Servomechanism
Let us start with the negative feedback loops. Wiener's (1961) servomechanism, which has been ably developed by William Powers (1973). The basic idea is no doubt familiar (see Fig. 5). A comparator generates an error signal from the difference between the perceptual input and an internal reference signal. The error signal then selects an output function which, when enacted, will bring the system closer to its goal. Powers begins by stacking his servomechanisms (see Fig. 6), one level above the other, so that higher-level servos determine the reference signals for lower-level servos. His originality, however, lies in associating each level in the stack with a different order of experience, a different set of ontological types such as sensations (e.g. colour, tension, texture), configurations (e.g. bear, pine tree, standing), and sequences (e.g. trot, jump). Higher-order foci are abstracted from residuals passed on by lower-level systems (Benzon, 1978; Hays, 1981).
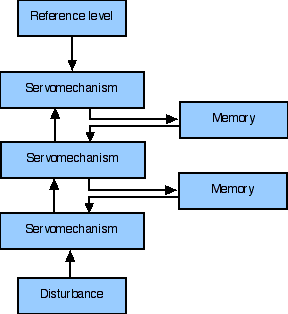
Fig. 6. A stack of servomechanisms
Returning to the diagram, you will notice, off to the side, a series of boxes labelled 'memory', with each box being connected to the servomechanical stack at a specific level. In Power's account, these boxes contain stored reference signals, schemas, for their associated servos. At the configuration level, we would have schemas for, e.g., particular postures, particular objects. At the sequence level we have schemas for motor, auditory and visual sequences. We find it natural to think of the brainstem and the peripheral nervous system as a stack of servos, while the cortex is a memory unit. The cortex stores schemas across all orders. It thus stores foci and residuals abstracted from the analysis of lower level states of activity (cf. Edelman, 1978; p. 71). It now needs a mechanism for choosing among stored schemas.
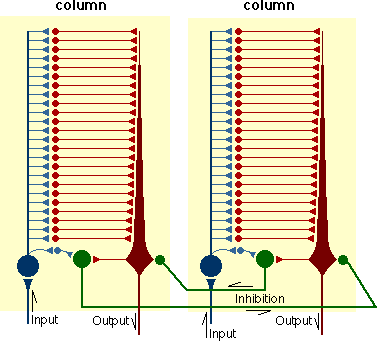
Fig. 7. OCCAM (adapted and redrawn from Spinelli, 1970)
Here, we propose a closed loop selection mechanism based on the positive feedback heterostatic model of the neuron developed by Harry Klopf (1981) and on Spinelli's OCCAM (1970: Metzler & Spinelli, 1977). In OCCAM (see Fig. 7), each cortical column is tuned to a pattern of inputs, which is stored by differential synaptic thresholds between input and output neurons (see also Longuet-Higgens, 1968)). Input to a cortical region is available to all columns. But only those whose stored pattern is a close match for the arriving input will be able to respond.
The local circuitry in the column is such that only an input which matches the stored pattern closely will propagate through to the central pyramidal output cell strongly enough to generate an output. The match cell compares the input with the output. When the match between the two is close, the match cell will fire and exert a strong inhibitory effect on neighbouring columns, acting to turn them off. If the match between input and output is weak, the inhibitory effect will be weak. Such a column is likely to be totally inhibited by other columns whose match is good. Each column thus acts like Klopf's (1981) heterostat; it acts to maximize its excitatory input. In such a device, a diminishing difference between stored value and input will result in increased output; the device is thus a positive feedback device. (In a negative feedback system, a diminishing difference between input and stored value will result in diminishing output.)
The overall effect is that schemas become stored in columns which are widely distributed across a given cortical region. These populations of columns are in competition with one another. The population whose schema most closely matches the input will be able to fire strongly and thereby inhibit firing in other populations. If the system becomes deadlocked, that is, if no stored schema is a significantly better match than any other schema, it is necessary to establish a new schema in the system to handle the input. Deadlock is thus a sign that the system must go into a mode for reorganization, or learning.
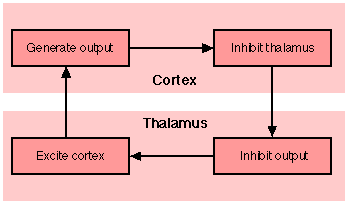
Fig. 8. Alpha wave generation. Note: These boxes indicate control functions, not discrete physical entities
From this general characterization, we can derive an account of alpha wave activity (Fig. 8). We know that corticofugal stimulation has an inhibitory effect on the thalamus (Shepherd, 1974). The cortical efferents are outputs from heterostatic populations of columns, while inputs to those columns come from thalamic nuclei. The cortex is thus acting to inhibit its sources of input. Since, however, the cortex consists of closed loop devices which are driven by their inputs (note that the cortex has no spontaneous electrical activity; Milner, 1976; p. 51), the inhibition of thalamico-cortical efferents by the cortex will dampen the activity of cortical neurons generating output to the thalamus. With cortical inhibition released, the thalamic neurons can once again generate output to the cortex, which then inhibits the thalamus. We thus have a rhythmic cycle, suggesting a mechanism underlying thalamico-cortical alpha waves (cf. Thatcher & John, 1977). When a cortical system successfully analyses its input, i.e. identifies the category (population of tuned cortical columns) to which it belongs, we get this rhythmic firing. When the cortex is unable to identify the input, when it is deadlocked, we will not get this rhythm. This is thus consistent with the observed relationship between high alpha output and the absence of novel stimuli (Pribram, 1971). Novel stimuli will not drive any population of columns strongly enough for them to set up a stable and rhythmic pattern of activity.
We can conclude the discussion of the decision principle by noting that its effective realization requires a diagonalization over the interaction between the positive feedback selectional system and the negative feedback goal-seeking system. It is, in fact, from this diagonalization that Powers' servomechanical orders achieve their coherence. It is this diagonalization which achieves the embedding of a sensation space within a configuration space (ordinary three-dimensional space), configuration space within space-time sequences and space-time sequences within casual interactions. This higher-level diagonalization is a positive-negative feedback stack (Fig. 9); it is the deepest implementation of the decision principle.
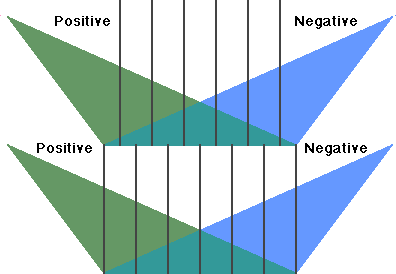
Fig. 9. A positive-negative feedback stack
Finitization: the figural principle
The figural principle concerns the relationship between Gestalt
or analogue process in neural schemas and propositional or digital processes. In
our view, both are necessary; the figural principle concerns the relationship between
the two types of process. The best way to begin is to consider Miriam Yevick's work
(1975, 1978) on the relationship between 'descriptive and holistic' (analogue) and
'recursive and ostensive' (digital) processes in representation.
The critical relationship is that between the complexity of the object and the complexity
of the representation needed to ensure specific identification. If the object is
simple, e.g. a square, a circle, a cross, a simple propositional schema will yield
a sharp identification, while a relatively complex Gestalt
schema will be required for an equivalently good identification (see Fig. 10). Conversely,
if the object is complex, e.g. a Chinese ideogram, a face, a relatively simple Gestalt
(Yevick used Fourier transforms) will yield a sharp identification, while an equivalently
precise propositional schema will be more complex than the object it represents.
Finally, we have those objects which fall in the middle region of Figure 10, objects that have no particularly simple description by either Gestalt
or propositional methods and instead require an interweaving of both. That interweaving
is the figural principle.
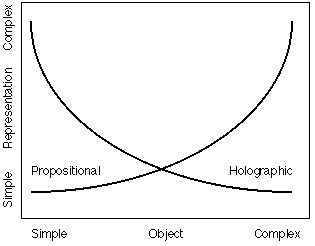
Fig. 10. Yevick's law. The curves indicate the level of representational complexity required for a good identification
Definition. The figural mechanism brings environments of moderate complexity within the limits of computability by putting a propositional assemblage of local narrow band-width Gestalts into a framework provided by global wide band-width analysis to achieve cross-validation of the two analyses.Let us begin by considering vision. It is well known that we have a very high capacity for single glance recognition of visual scenes and objects (Haber, 1970). Perhaps the most dramatic example is that of a child being able to recognize individual phonograph records from their global physiognomy, rather than reading the label (Church, 1966). The obvious mechanism for achieving this is a global transformation, such as the Fourier transformation. Such global processes do, however, have limitations, one of which is illustrated by what we call the 'strange friend' phenomenon. The phenomenon occurs when someone you know acquires or shaves off a beard, changes hair-style, etc. You recognize the person but you also sense that something has changed. You inspect the person and eventually you figure it out--or perhaps you do not, perhaps you have to be told what has happened. What is going on?
Between contextual clues, such as name, location (your friend's living room), voice, etc., and wide band-width information, the general shape of the head (Harmon, 1973), you have no trouble recognizing the person as your friend. But the change is significant enough that the current image of your friend no longer is a satisfactorily close match to your global schema for him or her; so you inspect the face until you have determined the change. This inspection, however, requires the use of a propositional representation. On the basis of eye movement data, Noton & Stark (1971) have proposed such a representation, which they call a feature ring. The object is represented as a ring of features (see Fig. 11). A different feature is brought into focus with each fixation point, while the direction of the oculomotor path between two features encodes the propositional relation between them. We propose one change in this analysis. A feature, we suggest, is a relatively narrow band-width component in a local area of the object (cf. Pribram, 1981). Hence, the feature ring is part of a mechanism relating local narrow band-width Gestalts to a global wide band-width Gestalt . Positive identification requires that the two Gestalts , composite narrow band-width and global wide band-width, be cross-validated. If they fail to match, one or the other is suspect. Returning to the strange friend, the inspection of local areas of the friend's face allows one to find the local area, or areas, which register the poorest match with the internal schema, those areas where the positive feedback selection process fails to stabilize. These areas pinpoint the change.
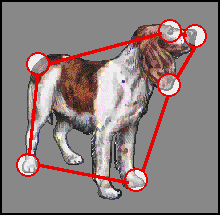
Fig. 11. A feature ring
Now let us consider a different phenomenon, cross-modal matching of objects in visual and haptic space. How do we determine that visual and certain haptic stimuli specify the same shape, e.g. circle, square, triangle? The task is difficult, and for a while it was believed that only humans could do it, but recent research has shown that monkeys and apes can also do it (Petrides & Iverson, 1976; Davenport & Pralinsky, 1977). The identification of objects in haptic space requires active palpation (Gibson, 1966; Werner, 1974). This suggests that haptic schemas involve the propositional integration of several haptic images where, as in the case of vision, the propositional relations are derived from the motor schemas which guide the sensors over the object. The visual and haptic triangles can be identified by the similar propositional relations which structure their scan paths. Thus, where diagonalization was sufficient to identify visual and kinesthetic motion (see 'Ontogeny'), the figural mechanism of propositional relations is needed to establish identity between visual and haptic shapes.
We can now consider possible neural implementation. Once again, consider the visual system. Figure 12 misses many structures, but it indicates the gross structure of the system, which is that the visual system is dual (Weiskrantz, 1974; Jones, 1974; Salapatek, 1975). Input from the retina goes to the superior colliculus and thence to the infero-temporal (IT) cortex by way of the pulvinar. In the other path, retinal input goes to the lateral geniculate nucleus (LGN) and then to the occipital cortex. The next level of processing occurs in the foveal prestriate cortex and then, finally, the IT cortex. Both the superior colliculus and the IT cortex are linked to the frontal eye fields. There is also an unindicated link between the prestriate cortex and the frontal eye fields.
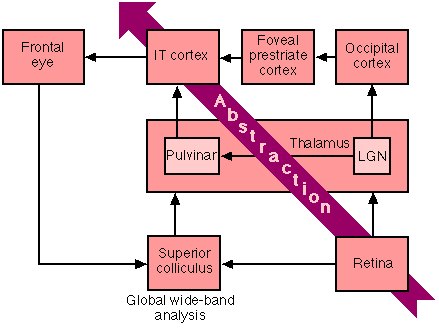
Fig. 12. The figural system
Let us begin with the superior colliculus (Gordon, 1972). It is organized into several layers, each being retinotopically organized. The receptive fields of cells near the surface are rather small, while those of cells in deeper layers are larger (and become multi-modal). The superior colliculus thus seems designed to conduct a crude, wide-band spectral analysis of the visual field in which the surface layers extract relatively narrow band-width information, while deeper layers extract wide band-width information. The collicular analysis should be sufficient to locate and follow objects (Humphrey, 1970), and to generate scan paths based on major visual gradients.
The occipital cortex, however, seems designed to extract narrow band-width information from relatively small portions of the visual field, no larger than 5 degrees (Pollen & Taylor, 1974; Pribram, 1981). These narrow-band perceptual atoms are then integrated into larger narrow-band Gestalts of whole objects (see Jonides, Irwin & Yantis, 1982) in the foveal prestriate cortex (Luria, 1973; Gross, 1973). This information is then passed on to the IT cortex where it is integrated with global wide-band information from the superior colliculus through the pulvinar.
Receptive fields in the IT cortex can be quite large, suggesting input from the superior colliculus, or a wide region of the occipital cortex, or both (Gross, Bender & Rocha-Miranda, 1974), and one famous cell responded most strongly to an image of a monkey's paw. This complexity also suggests integration of both wide-band (from the superior colliculus) and narrow-band (occipital) information. Further, when the pulvinar is destroyed, eliminating the global wide band-width analysis, IT neurons become sensitive to stimuli anywhere in the visual field, whereas their sensitivity is more localized when there is collicular input (Gross et al ., 1974). This suggests that the wide-band analysis divides the visual field into figure and ground, and the narrow band occipital analysis is used to identify the figure.
The IT cortex and foveal prestriate cortex are also known to be involved in saccadic scanning (Aschoff, 1974). The frontal eye fields are also involved in saccades, but they are active after the saccade has started, not before. This would suggest that the retinal-IT cortex system is capable of generating a sequence of targets for the saccades, one after the other. But the set of vectors connecting those targets, and relations between them, is a higher-order function which is stored in the frontal eye fields. This set of relations is propositional reconstruction of the image. Since it is located in motor cortex, it would be a plausible source of information needed to generate the complex programs needed to draw images (Benzon, 1982).
This analysis of the visual system is certainly a long way from being complete, but it does suggest how the figural principle could be implemented in neural structure in a relatively straightforward way. The important point is that the nervous system needs both Gestalt and propositional mechanisms for a subtle account of the world. The controversy between advocates of images and advocates of propositions (Kosslyn et al ., 1979; Ullman, 1980) thus seems to us mistaken; both types of process are needed.
Analysis: indexing
The indexing principle is about computational geometry, by which we mean the geometry,
that is, the architecture (Pylyshyn, 1980) of computation rather than computing geometrical
structures. While the other four principles can be construed as being principles of computation, only the indexing principle deals with computing in the sense
it has had since the advent of the stored program digital computer. Indexed computation
requires (1) an alphabet of symbols and (2) relations over places, where tokens of
the alphabet exist at the various places in the system. The alphabet of symbols encodes
the contents of the calculation while the relations over places, i.e. addresses,
provide the means of manipulating alphabet tokens in carrying out the computation.
The token at a place is a value and the place is identifiable by way of the relation
given an address (see Fig. 13). Thus, the structure of the computational space can
be used to locate various content items--that is, indexing. The possibilities of
indexed computing become particularly exciting when one realizes, as von Neumann did, that
values and places can be encoded in the same alphabet, making it possible to introduce
the manipulation of computational geometry into the content of computation.
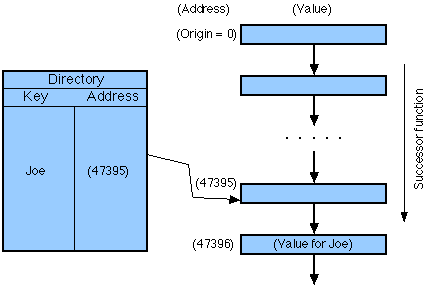
Fig. 13. Indexing
Within the context of natural intelligence, indexing is embodied in language. Linguists talk of duality of patterning (Hockett, 1960), the fact that language patterns both sounds and sense. The system which patterns sound is used to index the system which patterns sense. The names of concepts, the signifiers, to use Saussure's (1959) term, index those concepts. Linguistic concepts fall into two broad classes, contentives (nouns, verbs, modifiers) and functors (determiners, conjunctions, pronouns, prepositions, etc.). Loosely speaking, syntax orders functors and contentives so that the functors are relations over the places occupied by the contentives. Of course, the functors themselves occupy places in the system, and those places are addressed by the appropriate signifiers (see Bloom & Hays, 1978).
Definition. Indexing gives computational geometry, and language enables the system to operate on its own geometry.In order adequately to understand linguistic indexing, it is best to begin with a discussion of its cognitive underpinnings (Piaget, 1962; Vygotsky, 1962; Neisser, 1976). Think of the linguistic sign as a component in a multi-modal schema. Thus, the child's 'doggie' is visual, haptic, olfactory, motoric (the child can imitate the dog), general auditory (the barks, whines and growls the dog makes), and also linguistic auditory. 'Spot', the word 'doggie', and linguistic motor, the respiratory and articulatory gestures needed to utter 'Spot' and 'doggie'. Initially, the child treats the linguistic schemas just as attributes of the multi-modal dog. Eventually, however, the linguistic use becomes differentiated from the rest, for the linguistic attributes are the only ones directly under the child's control. The child can utter 'doggie' or 'Spot' at will, and the linguistic auditory attribute will invariably appear in the auditory channel.
But, while the child can explore the environment to make the visual, haptic, olfactory, and general auditory attributes available, the success of such activity depends, to some extent, on the dog's co-operation. Thus, the child will come to conceive of the relation between the linguistic attributes of the multi-modal dog schema and the non-linguistic attributes as being fundamentally different. With the advent of that distinction, that multi-modal schema has become differentiated into the linguistic signifier and the conceptual signified. The signifier is implemented in Broca's and Wernicke's areas (Lenneberg, 1967), but the crucial tissue is that which regulates the relation between those areas and the general multi-modal signified. This is parietal tissue Geschwind (1965) has found to be so crucial.
In this interpretation, motor control is central (see also Pribram, 1971). This emphasis supported by recent studies of cerebral blood flow (Lassen et al ., 1978, Orgogozo & Larsen, 1979), which show that the supplementary motor area is active during speaking and reading aloud. This suggests to us that the frontal cortex implements language at least in part by storing schemas for the relations over addresses, syntax roughly speaking--an interpretation consistent with Roman Jakobson's (1971) work on aphasia and Karl Pribram's (1971) emphasis on the frontal cortex as subserving context-sensitive processes. Given a concept A and its address, an indexed system moves to another address having a specified relationship (e.g. sequence, descended in a tree) to A's and returns the concept B stored at that address. We conclude that Wernicke's and Broca's areas implement the auditory and motoric address space which language provides, while the relations are implemented in the frontal cortex.
Now we are in a position to consider the general neural significance of indexing. Left hemisphere specializations for language, Broca's and Wernicke's areas, implement the addresses, while the directory linking the addresses to values is in parietal tissue (Geschwind, 1965; Luria, 1973). This directory has no intrinsic order, unlike the directory of the digital computer, which is a binary realization of the integers; but that is a secondary matter. We do, however, have to consider the general issue of hemisphere specialization. Bradshaw & Nettleton (1981) have reviewed much of this material and find that while a general distinction between Gestalt , holistic, or spatial (right hemisphere, and linguistic or analytic (left hemisphere) processing can be sustained, it should not be interpreted as an exclusive dichotomy, but rather as two tendencies along a continuum. They go on to emphasize that another contrast is perhaps even more fundamental. This is a contrast between tasks involving duration, sequencing, temporal order and rhythm on the left, and on the right tasks involving exteroceptive body space, and the positions of fingers and limbs in relation to target positions.
The left hemisphere specialization for sequential processes is consistent with our analysis of the requirements of the indexing principle. The process of moving through an indexed computational space always involves sequential processing (in the digital computer this is the familiar fetch-execute cycle). In view of the blood flow evidence and general characterization of frontal versus anterior cortical function (Pribram, 1971; Luria, 1973), the left frontal cortex seems particularly important. In this view, the right hemisphere is not deeply indexed. Thus, it is not surprising that the speech which is mediated by the right hemisphere is retrieved in Gestalt units (Bradshaw & Nettleton, 1981) and is stronger in comprehension than production. This suggests that signifier and signified are relatively undifferentiated in the right, hence indexing is weak. Thus, right hemisphere superiority may primarily involve figural processes at the highest level. There is, however, a right hemisphere specialization which suggests a way of exorcising the ghost of the transcendental self, which Gunther Stent (1975) finds blocking the way to a scientific understanding of human behaviour.
For the right hemisphere does appear to be specialized in the representation of the self-image (Luria, 1973). Damage to the right parietal region will leave the patient without any sense of loss, while similar damage on the left will not similarly go unnoticed. Self-image thus seems to be in the right hemisphere. But that self-image is linguistically indexed in the left hemisphere. The process of manipulating and operating on that self-image by the use of linguistic indexing is, in our interpretation, the core of our senses of self. The most obvious behavioural manifestation of this is the self-regulating function of speech investigated by Vygotsky (1962) and Luria (1959). We gain control over our actions by talking to ourselves, initially aloud; but later through silent, inner speech. The body image may be on the right, but the left hemisphere systems indexing that image are not similarly imaged on the right. Those left hemisphere systems are invisible to the basic self-image; the self thus appears to have a transcendental and incorporeal aspect.
We conclude this section by noting that the indexing principle enters information into the ontology of the system. Not only does the system deal in modes, in ontological types, goals and categories, abstractions and relations, it now deals with information.
The other objects our nervous system presents to us seem to exist in the world; but informatic objects are elusive. They always have physical embodiment, but the physical substance of the embodiment is irrelevant. The information resides in the relations which are indexed by that physical substrate, not in the schemas which might be associatively linked to the substrate. And if we as theoreticians can see the entire system as an information processor, that is because our nervous system gives us the basic capacity, though language, to explicitly grasp and attend to information, rather than merely to be information. The principle by which an information processing system presents itself with informatic objects, rather than using information to present non-informatic objects, is the indexing principle.
Ordering the principles
The five principles have an intrinsic ordering such that the implementation of a principle late in the ordering presupposes the implementation of principles early in the ordering. We have explicated the principles in intrinsic order. We now justify that order and introduce a set of terms from Hays (1981; Hays & Benzon, 1981) and Benzon (1978) which can be used to designate perceptual and cognitive systems according to their cumulative implementation of the principles of intelligence.
Sensorimotor intelligence
Diagonalization presupposes mode. Modal control is the basic principle. By linking conditions in the organism's inner and outer environments together into distinct behavioural modes, the modal principle stabilizes the relationship between the system and the external environment so that the system is not continually trapped into the vertiginous flutter which afflicted the legendary ass equidistant between two bales of hay, or, in the general case, the organism deciding among more or less equally plausible goals (see also Powers, 1973; 253 ff.). Modality further gives the system control over its own implementation. With the relationship between inner and outer worlds stabilized, and the foundation of self-control laid, diagonalization in the external world becomes possible.
Systemic intelligence
Action presupposes diagonalization. Diagonalization provides the organism with a stable system of ontological types; the organism now lives in a world of objects rather than a world of features and gradients. It now becomes feasible to fix categories within the ontological types and to link those categories to a hierarchy of control system goals. In such a system, diagonalization embeds ontological types into a hierarchy according to the structure of the goal hierarchy, property space within geometrical space, geometrical space within the space-time, space-time within cause, etc.
Episode intelligence
Figural composition and abstraction presuppose diagonalization. Propositional is an active, a fundamentally motoric, process. The mechanism which traces propositional paths and thereby composes a narrow band-width schema (or, from another point of view, decomposes the global Gestalt into component regions) must be of systemic level, an action system. Diagonalization through schemas at this level identifies propositional and Gestalt accounts of the same object, thus yielding an abstraction.
Gnomonic intelligence
Indexed sensory analysis and motor programming presupposes figurality. A gnomonic system requires an episodic system just to manage the indexing function, regardless of whether or not any of the schemas indexed are of episodic degree. Diagonalizing in an indexing system adds informatic objects to the system's ontology. It recognizes language as being distinctly different from anything else in its world.
With the order of the principles thus fixed, the direction of evolution is fixed. The crucial issue then concerns the conditions which support and even necessitate this evolution.
The development of natural intelligence
Given the five principles and their intrinsic ordering, the problem of providing an
evolutionary account becomes one of identifying the environmental patterns which
move evolution from one principle to the next and of identifying the neural structures
which implement this movement. This much is a descriptive problem, one which we undertake
in the next section ('Phylogeny'), to be followed in 'Ontology' by some remarks about
ontogeny, especially the human case. The purpose of this section is thus to display the relationship between the informatic principles and biological principles.
However, before we undertake the descriptive task, we want to point out why it is
not explanatory; it is an account of how the nervous system evolved, principle by
principle. But it does not explain why. To do that would require an extension into
the physical realm where the concept of entropy can be used to commeasure informatic and
energetic costs and benefits (Ashby, 1956; Wiener, 1961; Tribus & McIrvine, 1971).
The general direction of such an explanation would be to show that the energetic
costs of new information processing are low in relation to the energetic benefits, but making
such an explanation would take us too far from the main line of our exposition.
Phylogeny
Starting with a primitive vertebrate (see Table 1), we sketch an evolutionary sequence modelled after Jerison's (1976) account of paleoneurology. The general idea is to suggest the peripheral receptors and effectors and the central integrating mechanisms which make movement to an informatically richer niche possible.The primitive vertebrate nervous system is reticular (Best, 1972; Bowsher, 1973). The only principle active at this level is the modal principle. Within a given mode, behavior is governed by on-blocks, where the conditional elements are innate releasing mechanisms and the executed programs are fixed action patterns (Lorenz, 1969). These on-blocks are executed as they are triggered by the interaction of environmental stimuli and organismic modal shifts. There is little or no autonomous chaining of these on-blocks.
Diagonalization is promoted by the move to land, although perhaps not initially, since early amphibians are no more encephalized than fishes (Jerison, 1976). Terrestrial environments are more demanding than aquatic ones. Gravity is a harsher gradient, the visual world is more complex, and so are the auditory and olfactory worlds. In such a world, being able to deal with objects, as opposed to mere features and gradients, would have survival value.
Some diagonalizations may be genetically determined, while others are learned. For example, Eibl-Eibesfeldt (1979) reports that newly metamorphosed frogs will snap at anything that moves, including leaves and pebbles; but they quickly learn to avoid such harmful prey. The developing cortex is where the diagonalization takes place (recall the discussion of tilt constancy in 'Coherence: diagonalization').
However, the organism must have available a mode in which it accomplishes the diagonalization. We can call this mode reorganization (Powers, 1973). There is experimental evidence that reptiles are better than fish at habit-reversal conditioning regimes (Bitterman, 1965). In such a regime, the organism is first conditioned to stimulus A and then, when the preference is strong, conditioned to B, and then to A, etc. Turtles improve in this situation (i.e. the number of trials between reversals diminishes) but fish do not. They are thus better equipped to survive in a complex environment where objects may change their significance than is the fish.
The evolution of mammals gives us the eventual emergence of the action principle. According to Jerison (1976), mammals were initially relatively small animals inhabiting nocturnal niches left free by the then dominant dinosaurs. Peripherally, they developed smell and hearing. Later, when extinction of reptiles made daytime niches available, they developed the mammalian visual system with its rich thalamic and cortical structures. Along with this increased variety in sensory systems comes increased specificity in motor control through the corticobulbar and corticospinal tracts, which provide direct cortical connections with motor neurons (Ghez, 1981). Thus, the mammalian nervous system has a layer of high resolution sensorimotor system around a reptilian core of lower resolution systems. The expanding neocortex implements the central perceptual and cognitive aspect of the mammalian systems and diagonalizes through them to create the mammalian world of space-time (Jerison, 1976), in which the decision principles operates. At this point in evolution, the neocortex begins to differentiate into distinct sensory and motor regions (Sarnat & Netsky, 1974), which would be needed to support the more sophisticated diagonalization which integrates these new structures into a coherent system.
The behavioural mode which is most important here is play (Smith, 1982), which we are interpreting as a general mode of behaviour (Schwartzman, 1982) which can be adopted toward any of the activities the organism executes. The complexity of the behavioural repertoire has now gone far beyond that which can be genetically specified or 'wired' to the environment through simple imprinting mechanisms. Now a great deal of experience is necessary. Play is the mode in which that experience is gained without becoming rigidly fixed before the environment has been adequately explored.
The most sophisticated mammals are, of course, primates, whose arboreal existence requires manipulative capacity and good depth perception. The evolution of stereoscopic vision (Sarnat & Netsky, 1974) provides the needed depth perception and the further development of the corticospinal system, especially the large Betz cells which synapse directly on spinal motor neurons (Evarts, 1979; Ghez, 1981), provides the needed manipulative capacity. The central control of manipulation in stereoptic visual space evolves into the figural principle, with the enlargement of association cortex being the means of implementing this principle.
The behavioural mode appropriate to this level of organizations is visuo-motor imitation (Jolly, 1972), as opposed to the auditory-vocal imitation of some song birds (Hinde, 1972). While this certainly presupposes the diagonalization between motoric and visual space discussed in 'Coherence: diagonalization', it goes beyond that to require the propositional reconstruction of the recognized visual Gestalt into a sequence of motor impulses which will result in the successful reproduction of the observed behaviour. A most striking example of the role of imitation in primate life is male sexual behaviour, where imitative play in youth is essential to successful male mating (Ford & Beach, 1951). A primate male reared in isolation will be unable to mate when placed with a receptive female. Infra-primate males raised in comparable circumstances are still able successfully to mate.
Of primates, Homo sapiens sapiens , of course, has the most sophisticated cognitive system. The only peripheral elaboration which seems unique to man is the existence of monosynaptic corticomotor neurons innervating respiratory muscles, which would be necessary for breath control in speech (Phillips & Porter, 1977). Otherwise, the elaborations are central, Broca's and Wernicke's areas, hemisphere specialization. While we do have an idea of how the linguistically specialized cortical areas might have come about, we reserve that discussion for the next section, concerning ontogeny. Our immediate objective is to provide a fairly standard type of evolutionary sketch.
The final push in human evolution came about 30,000-50,000 years ago during the late Pleistocene era, but it was preceded by a period in which a variety of hominid forms emerged which show evidence of tool use and upright walking (Pilbeam, 1972). In our view, tool use developed during this period through the elaboration of a visuo-manipulative figural system employing imitative learning (cf. Wright, 1978). If we attribute an increase in social complexity to these precursors, as Vernon Reynolds (1976; cf. Lovejoy, 1981) does, that might well have been subserved by an auditory-vocal call system. The final step taken in the late Pleistocene would be to use the auditory-vocal call system to index the visuo-manipulative tool system. The initial development of these two systems would have been subserved by hemisphere specialization and the inter-relation facilitated by the emergence of vocal and auditory cortical tissue specialized for language, permitting the creation of a separate indexing space. The new behavioural mode is, of course, the linguistic mode. It may seem strange to regard language as a mode, or perhaps as a set of modes, but the cerebral blood flow evidence points in this direction and reinforces our general sense that language involves humankind in an essentially new type of relationship to self and to the world.
Ontogeny
The genes must build the nervous system step by little step. Any serious consideration of those mechanisms is certainly outside the scope of this paper, for those processes are deep within the realm of biological control systems, but we do want to make some remarks about the ontogeny of these principles in man.We can set the stage with Gerald Edelman's (1978) group selection theory of cortical function. Edelman assumes that, when they mature, cortical columns already prefer certain inputs over others. These preferences need not be finely tuned to the environment, but there should be a large repertoire of them so that columns will respond differentially when exposed to external inputs. When exposed to an environment, different sets of columns would be attracted to different stimuli, would come to 'store' the neural patterns set up by those stimuli (see also Spinelli, 1970). The repertoire of selective preferences for the early maturing columns could be determined by embryological processes, while the repertoires of later maturing columns could be guided by gradients set up in the neural tissue by the interaction between the already mature areas of the brain and the external world.
It is useful to remember that, while the cerebral cortex appears to be a complex three-dimensional object, it is in fact a flat sheet which has been crumpled up to fit into the skull. This sheet matures from the edge to the centre, and, in humans, it matures post-natally (Milner, 1976), with there being enough growth in the third decade that the sutures of the human skull do not set until the late twenties (Gould, 1977). Consequently, much of the basic 'wiring' of the cortex is under environmental guidance. This is important, for there really is no other way to explain, for example, Gunnar Johansson's results (see 'Coherence: diagonalization'). That it took subjects only about a tenth of a second to identify the moving dots as walking and dancing people suggests that the identification process is 'hard-wired'. It is too much to expect the genetic code to accomplish such 'wiring' without any guidance from the environment. But, if the 'wiring' was not completed until environmental exposure was available, then, we hypothesize, the problem is not so intractable (for the genes, not necessarily for developmentalists), environmental input can guide the final 'wiring'.
Human brain maturation is not even, it occurs in stages (Luria, 1973, p. 87; Epstein, 1974). These growth stages correspond well to the general stages Piaget has postulated (in, e.g., Piaget & Inhelder, 1969), and we find it attractive to match these stages with the successive maturation of the brain centres which implement each of the informatic principles. Figure 14 sketches out the relationship between brain maturation (based on Lenneberg, 1967; Lancaster, 1968; Milner, 1976; Parker & Gibson, 1979) and behavioural growth. It is interesting that alpha wave production increases in stages (Epstein, 1980), from zero EEG energy at birth to about 70% in adults. Given the explication of alpha in 'Action; the decision principle', where it is seen to derive from the successful functioning of the positive feedback selective mechanism, these spurts in alpha energy would indicate the maturation of tissue capable to diagonalizing over successively more complex patterns of residuals. Thus, the leveling point in each of the maturation curves of Figure 14 comes at one of these alpha spurts. The first curve turns when the modal mechanism is well enough established that diagonalization becomes possible (recall 'Ordering the principles'). The second curve, the diagonalization curve, turning when the mechanisms of diagonalization are well established, gives rise to the Piagetian world of constant objects. Each successive curve turns when diagonalization over the correlative principle becomes solidified. While it is not possible to give any detailed discussion of this concept of the relationship between neural and behavioural ontogeny, we do want to touch lightly on the role of language.
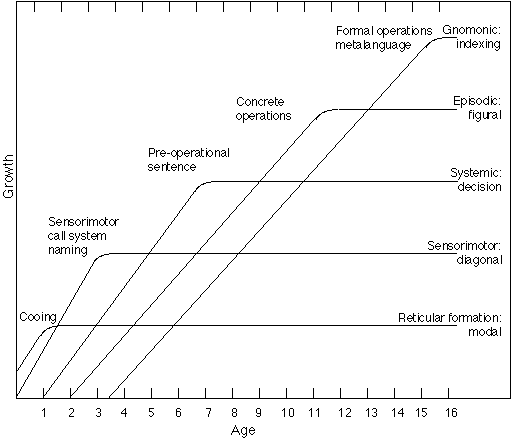
Fig. 14. Ontogeny
First, let us note that human ontogenesis is not simply an extrapolation of monkey or ape ontogenesis; it has a shape unique to man (Lenneberg, 1967). As an adaptation to large brain size and the relatively small pelvic girdle of bipedal mothers, the infant is born unusually early in development so that approximately the first year of life is spent, in effect, as an extra-uterine embryo (Campbell, 1966). At the other end of development, the sutures in the human skull close well after sexual maturity, whereas those in ape skulls close before sexual maturity (Gould, 1977). We want to suggest that the relatively early birth of humans makes language possible and that language makes late closure of the skull necessary.
During the first year, the infant babbles with increasing approximation to the sounds of the parental language. This is largely under subcortical control, but it provides the gradients necessary for some primary motor and auditory tissue eventually to become specialized for language input and output (Broca's and Wernicke's areas). This early availability of tissue diagonalized for the development of a vocal-auditory channel is crucial if language is to become a major medium of communication.
Apes do not have well-articulated cortical control over vocalization (Myers, 1978), nor does vocalization have the central role in ape communication that it has in human communication (Jolly, 1972). We link this to a difference in the maturation of attachment, which is central to primate social life (Bowlby, 1969, Rajecki, Lamb & Obmascher, 1978). Among apes, attachment behaviour starts almost immediately after birth (Bowlby, 1969). Comparable human behaviour is delayed about 6 months. Thus, by the time a human infant is ready to begin a genuinely social life the neural basis for a vocal-auditory communication channel has been laid; there is nothing comparable in ape ontogeny.
It is thus natural for humans to incorporate speech into their social life. It would be unnatural for infra-human primates to do so. They do not have a vocal-auditory channel available to them when they being attachment, and once attachment has established the basis for social life it is probably impossible to incorporate a genuinely new communication channel into it; a critical period will have passed, the tissue is set. Thus, a crucial step in the development of indexed cognition and communication is early exposure to the world leading to the creation of a linguistic channel before or concomitant with the origins of attachment behaviour.
This suggests, as Stephen Jay Gould has argued informally (1979), that the emergence of bipedal walking was the crucial seed driving human evolution. Bipedalism necessitated early birth and early birth favoured the development of a vocal-auditory interaction channel in the infant before attachment began. The vocal-auditory system can then be used to index the visual-manipulative tool system (recall 'Analysis: indexing', 'Phylogeny').
Given the emergence of babbling and cooing, we suggest that the further development of language involves a reconstruction of the diagonalization, action and figural principles within an indexed computational system. This is, however, only pseudo-indexing. The distinction between words and things is notoriously difficult for children (Piaget, 1962; Vygotsky, 1962). Full control of the indexing demands the maturity of tertiary cortical tissue, which is the last to mature (Lancaster, 1968). Until that tissue is fully mature, the child is dependent upon interaction with adults to preserve the integrity of his or her conceptual system.
Referring to Figure 14, the sensorimotor development of vocal and auditory tissue diagonalized for speech provides the necessary foundation. The basic requirement of indexing, duality of patterning, has been met. Now it is possible to reconstruct each of the principles within an indexed computational space. The reconstruction of diagonalization within an indexed space yields elementary semantics, the linking of signifier to signified (see Bloom & Hays, 1978). The child is now capable of holophrastic utterance (Lenneberg, 1967), single morphemes which designate whole propositions in an undifferentiated way.
With the beginning of single two-word utterances, such as 'here sock', 'more milk', syntax begins to emerge. Syntax is the reconstruction of the decision principle within an indexed computational space. Thus conceived, syntax is a mechanism for selecting concepts and combining them into strings which meet the immediate goals of communication.
The reconstruction of figurality yields the capacity to generate a coherent discourse (see Phillips, 1978). The global wide-band Gestalt of figurality corresponds to the intuitively sensed theme of the discourse. The discourse proceeds though the propositional generation of a succession of narrow-band local Gestalt , each of which is rendered into a phrase or clause through the syntactic action principle.
When the gnomonic capacity explicitly to regulate indexing matures, we have the emergence of Piaget's formal operations period. Now the child is capable of reliably regulating the relationship between word and thing, and can thus have concepts about information, i.e. concepts about language and thought. For an essential function of language is its metalingual capacity to focus attention on itself (Jakobson, 1960).
Within this realm, abstract ideas emerge from discourse through the mechanism of metalingual definition (Hays, 1973, 1976, 1981; White, 1975; Benzon, 1976, 1978, 1981; Phillips, 1978; Hays & Benzon, 1981). Metalingual definition uses a story to define a term; the term designates the wide-band global Gestalt over the story (see Fig. 15). Thus, charity is when someone does something nice for someone else without thought of reward. Charity is abstractly defined by a generalized story (is when . . .). The story consists of narrow-band Gestalts , and any pattern of such Gestalts which matches that associated with charity is deemed to be a story about a charitable act. The technique of metalingual definition is quite flexible and general and can be used recursively to define higher-level abstractions over stories containing lower-level abstractions.
While the cortical mechanisms of metalingual definition are not clear enough to us that we are willing to hazard a diagram analogous to the visual figural diagram (Fig. 12), it is clear to us that many cortical areas are involved. Penfield & Roberts (1959) describe regions in the temporal cortex where stimulation yields experiential responses, the patient re-experiences episodes from the past. Thus, we have the stories which are the basis of metalingually defined concepts; the individual episodes of the stories are the narrow-band atoms. In roughly the same region, Penfield & Roberts also report interpretative responses, the patient feels that the operation has happened before, or that the current situation is absurd, or distances are altered. These areas seem to encode high-order perceptual parameters (Benzon, 1978; Hays, 1981) which govern the interpretation of entire scenes. They are parameters which apply to informatic objects, which enter into the system with the advent of indexing ('Analysis; indexing'). It is the scene which is familiar or strange, near or far. The frontal lobes are also essential, for they are intimately involved in planning, the linear structure of narrative, and analysis (Luria, 1973). In general, the frontal lobes store the relations which link atoms, whether figural or indexical, together into sequences (recall 'Finitization: the figural principle' and 'Analysis: indexing'). When we then add in the linguistic regions of the cortex and the multi-modal parietal zones (Luria, 1973; Lynch, 1980), we find ourselves covering the cortex. Metalingual definition is thus a mechanism for integrating the activity of the higher cortical regions which allows the brain to identify abstract patterns of objects and events in the world.
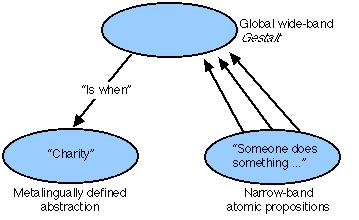
Fig. 15. Metalingual definition
Now we return to human ontogeny and the question of just how it is that language necessitates brain growth extended beyond sexual maturity. If we assume that the skull sutures set when the hypothalamus, which controls growth (Tanner, 1970), in some way senses that the brain is no longer growing, we need only suggest why the emergence of language stimulates brain growth. With the emergence of the linguistic capacity of metalingual definition, the human cognitive system is no longer closed. Abstractions can be created and learned indefinitely; but doing so requires the extended availability of tissue in which the abstractions can be implemented. Consequently, the brain continues to grow in order to accommodate the demands of this new, this emergent, cognitive capacity. With the emergence of full indexing capacity, the brain is no longer restricted to the world immediately given by the various sensory channels and the informatic principles. For the inclusion of the final principle, indexing, allows the implementation of a new cognitive structure which is essentially free of the conditions set by the earlier principles.
Thus, the ontogenetic sequence which eventuates in human language differs from that of apes at both its beginning and its end. Early exposure of the environment lays the groundwork for indexing. But indexing is not fully operative until the final maturation of tertiary cortical tissue implements reliable regulation of the relationship between signifiers and signifieds. And the ultimate effect of this is to make such a rich conceptual world available that brain growth continues on beyond sexual maturity.
References
Aschoff, J.C. (1974). In (F.O. Schmitt & F.G. Worden, Eds): The Neurosciences Third Study Program.
Cambridge, Massachusetts: MIT Press, pp. 305-310.
Ashby, W.R. (1956). An Introduction to Cybernetics.
London: Chapman & Hall Ltd.
Benzon, W. L. (1976). MLN
91
, 952-982.
Benzon, W. L. (1978). Cognitive science and literary theory
. Ph. D. dissertation, Department of English, State University of New York at Buffalo.
Ann Arbor University Microfilms 78-10602.
Benzon, W. L. (1981). Lang. Style
14
, 251-270.
Benzon, W. L. (1982). In (M. Herzfeld & M. Lenhart, Eds): Semiotics 1980.
New York: Plenum Press, pp. 27-36.
Bernstein, N. A. (1967). The Coordination and Regulation of Movement.
London: Pergamon.
Best, J. B. (1972). In (A. D. Breck & W. Yourgrau, Eds): Biology, History, and Natural Philosophy.
New York: Plenum, pp. 37-78.
Bitterman, M. E. (1965). In (J. L. McGaugh; N. M. Weinberger & R. E. Whalen, Eds):
Psycho-biology.
San Francisco: W. H. Freeman (1967), pp. 150-158.
Bloom, D. & Hays, D. G. (1978). In (J. Hinds. Eds): Anaphora in Discourse.
Edmonton, Canada Linguistic Research, Inc., pp. 1-68.
Bloom, F. E. (1981). Sci. Am.
245
(4), 148-169.
Bowlby, J. (1969). Attachment.
New York: Basic Books.
Bowsher, D. (1973). Brain Behav. Evol.
8
, 386-396.
Bradshaw, J. L. & Nettleton, N. C. (1981). Behav. Brain Sci.
4
, 51-91.
Campbell, B. G. (1966). Human Evolution.
Chicago: Aldine.
Chomsky, N. (1980). Behav. Brain Sci.
3
, 1-61.
Church, J. (1966). Language and the Discovery of Reality.
New York: Vintage.
Davenport, R. K. & Pralinsky, K. J. (1977). In (G. H. Bourne, Eds): Progress in Ape Research.
New York: Academic Press, pp. 251-258.
Dismukes, R. K. (1979). Behav. Brain Sci.
2
, 409-448.
Eccles, J. C. (1973). The Understanding of the Brain.
New York: McGraw-Hill.
Edelman, G. M. (1978). In (G. M. Edelman & V. B. Mountcastle, Eds): The Mindful Brain.
Cambridge, Massachusetts: MIT Press, pp. 51-100.
Edelman, G. M. & Mountcastle, V. B. (1978). The Mindful Brain
. Cambridge, Massachusetts: MIT Press.
Eibl-Eibesfeldt, I. (1979). Behav. Brain Sci.
2
. 1-57.
Epstein, H. T. (1974). Dev. Psychobiol.
7
, 207-224.
Epstein, H. T. (1980). Dev. Psychobiol.
13
, 629-631.
Evarts, E. V. (1979). Sci. Am.
241
(3), 164-179.
Fine, M. L., Keefer, D. A. & Leichnetz, G. R. (1982). Science
215
, 1265-1267.
Ford, C. S. & Beach, F. A. (1951). Patterns of Sexual Behaviour.
New York: Harper & Row.
Gallistel, C. R. (1980). The Organization of Action: A New Synthesis.
Hillsdale, New Jersey: Lawrence Erlbaum Associates.
Geschwind, N. (1965). Brain
88
, 237-294, 585-644.
Ghez, C. (1981). In (E. R. Kandel & J. H. Schwartz, Eds): Principles of Neural Science.
New York: Elsevier/North Holland, pp. 271-283.
Gibson, J. J. (1966). The Senses Considered as a Perceptual System.
Boston: Houghton Mifflin.
Gordon, B. (1972). Sci. Am.
227
(6), 72-82.
Gould, S. J. (1977). Ontogeny and Phylogeny.
Cambridge: Harvard University Press.
Gould, S. J. (1979). Nat. Hist.
88
, 40-44.
Gregory, R. L. (1966). Eye and Brain.
New York: McGraw-Hill.
Gross, C. G. (1973). In (E. Stellar & J. M. Sprague, Eds): Progress in Physiological Psychology
, vol. 5, pp. 77-123.
Gross, C. G., Bender, D. B. & Rocha-Miranda, C. E. (1974). Interotemporal cortex,
a single-unit analysis. In (F. O. Schmitt & F. G. Worden, Eds): The Neurosciences Third Study Program.
Cambridge, Massachusetts: MIT Press, pp.229-238.
Haber, R. N. (1970). Sci. Am.
222
(5), 104-112.
Hall, A. R. (1963). From Galileo to Newton: 1630-1720.
New York: Harper & Row.
Harmon, L. D. (1973). Sci. Am.
229
(5), 70-82.
Hays, D. G. (1973). SIGLASH Newsletter
6
, 8-11.
Hays, D. G. (1976). In (R. F. Geyer & D. R. Schietzer, Eds): Theories of Alienation.
Leiden Martinus Nijhoff, pp. 169-187.
Hays, D. G. (1981). Cognitive Structures.
New Haven: Human Relations Area Files Press.
Hays, D. G. & Benzon, W. L. (1981). Metagram Software--A New Perspective on the Art of Computation.
Final Technical Report RADC-TR-81-118. Rome Air Development Center, Air Force Systems
Command, Griffiss Air Force Base, New York.
Hermes, H. & Markwald, W. (1974). In (H. Behnke, F. Bachmann, K. Fladt & W. Suss.
Eds): Fundamentals of Mathematics, vol. 1, Foundations of Mathematics The Real Number System
and Algebra.
Cambridge Massachusetts: MIT Press, pp. 3-88.
Hinde, R. A. (1972). Biological Bases of Human Social Behavior.
New York: McGraw-Hill.
Hockett, C. F. (1960). Sci. Am.
203
(3), 89-96.
Humphrey, N. K. (1970). Brain, Behav. Evol.
3
, 324-337.
Jakobson, R. (1960). In (T. A. Sebeok, Eds): Style in Language.
Cambridge, Massachusetts: MIT Press, pp. 350-377.
Jakobson, R. (1971). Studies on Child Language and Aphasia.
The Hague Mouton.
Jerison, H. J. (1976). Sci. Am.
234
, 90-101.
Johansson, G. (1973). Percept. Psychophys.
14
, 201-211.
Johansson, G. (1975). Sci. Am
. 232
(6), 76-88.
Jolly, A. (1973). The Evolution of Primate Behaviour.
New York: MacMillan.
Jones, E. G. (1974). In (F. O. Schmitt & f. G. Worden, Eds): The Neuroscience Third Study Program.
Cambridge, Massachusetts: MIT Press.
Jonides, J., Irwin, D. E. & Yantis, S. (1982). Science
215
, 192-194.
Kelley, D. B. (1980). Science
207
, 553-555.
Kilmer, W. L. McCulloch, W. S. & Blum, L. (1969). Int. J. Man-Mach Stud.
1
, 279-309.
Klopf, A. H. (1981). The Hedonistic Neuron: A Theory of Memory, Learning and Intelligence.
Washington, D.C.: Hemisphere Publishing Corp.
Kosslyn, S. M., Pinker, S., Smith, G. E. & Shwartz, S. P. (1979) Behav. Brain Sci.
2
, 535-581.
Krieger, D. T. & Liotta, A. S. (1979). Science
205
, 366-372.
Kuhn, T. S. (1962). The Structure of Scientific Revolutions.
Chicago: University of Chicago Press.
Lancaster, J. B. (1968). In (P. C. Jay, Eds): Primates: Studies in Adaptation and Variability.
New York: Holt, Rinehart & Winston, pp. 439-457.
Land, E. H. (1977). Sci. Am.
237
(6), 108-128.
Lassen, N. A., Ingvar, D. H. & Skinhoj, E. (1978). Sci. Am
. 239
(4), 62-71.
Lenneberg, E. H. (1967). The Biological Foundations of Language.
New York, John Wiley & Sons, Inc.
Lewis, M. E., Mishkin, M., Bragin, E., Brown, R. M., Pert, C. B. & Pert, A. (1981).
Science
211
, 1166-1169.
Longuet-Higgins, H. C. (1968). Proc. R. Soc., Ser. B.
171
, 327-334.
Lorenz, K. (1969). In (K. H. Pribram, Eds): On the Biology of Learning.
New York: Harcourt, Brace & World, pp. 13-93.
Lovejoy, C. O. (1981). Science
211
, 341-350.
Luria, A. R. (1959). Word
15
, 341-352, 453-464.
Luria, A. R. (1973). The Working Brain
. New York: Basic Books.
Lynch, J. C. (1980). Behav. Brain Sci.
3
, 485-534.
MacLean, P. D. (1978). In (S. L. Washburn & E. R. McCown, Eds): Human Evolution: Biosocial Perspectives.
Menlo Park, California: Benjamin/Cummings, pp. 33-58.
Metzler, J. & Spinelli, D. N. (1977). In (J. Metzler, Eds): Systems Neuroscience.
New York: Academic Press, pp. 25-54.
Milner, E. (1976). In (H. Whitaker & H. A. Whitaker, Eds): Studies in Neurolinguistics,
vol. 1, pp. 31-102.
Mountcastle, V. B. (1978). In (G. M. Edelman & Mountcastle, Eds): The Mindful Brain.
Cambridge, Massachusetts: MIT Press, pp. 7-50.
Myers, R. E. (1978). In (S. L. Washburn & E. R. McCown, Eds): Human Evolution: Biosocial Perspectives.
Menlo Park, California: Benjamin Cummings, pp. 59-76.
Nauta, W. J. H. & Feirtag, M. (1979). Sci. Am.
241
(3), 88-111.
Neisser, U. (1976). Cognition and Reality.
San Francisco: W. H. Freeman.
Noton, D. & Stark, L. (1971). Sci. Am.
224
(6), 34-43.
Olds, J. (1977). Drives and Reinforcements.
New York: Raven Press.
Orgogozo, J. M. & Larsen, B. (1979). Science
206
, 847-850.
Parker, S. T. & Gibson, K. R. (1979). Behav. Brain Sci.
2
, 367-407.
Penfield, W. & Roberts, L. (1959). Speech and Brain-Mechanisms.
Princeton University Press.
Petrides, M. & Iverson, S. D. (1976). Science
192
, 1023-1024.
Phelps, M. E., Kuhl, E. D. & Maziotta, J. C. (1981). Science
211
, 1445-1448.
Phillips, B. (1978). Am. J. Comp. Ling.
Microfiche 82.
Phillips, C. G. & Porter, R. (1977). Corticospinal Neurons: Their Role in Movement.
London: Academic Press.
Piaget, J. (1954). The Construction of Reality in the Child.
New York: Basic Books.
Piaget, J. (1962). Play, Dreams, and Imitation in Childhood.
New York: W. W. Norton.
Piaget, J. & Inhelder, B. (1969). The Psychology of the Child.
New York: Basic Books.
Pilbeam, D. (1972). The Ascent of Man.
New York: MacMillan.
Pollen, D.A. & Taylor, J. H. (1974). In (F. O. Schmitt & F.G. Worden, Eds): The Neuro-sciences: Third Study Program,
pp. 239-248.
Popper, K. (1959). The Logic of Scientific Discovery.
New York: Basic Books.
Powers, W.T. (1973). Behavior: The Control of Perception.
Chicago: Aldine.
Pribram, K. H. (1971). Languages of the Brain.
Englewood Cliffs, New Jersey: Prentice-Hall.
Pribram, K. H. (1981). Cognit. Brain Theory
4
, 105-122.
Pribram, K. H., Nuwer, M. & Baron, R. (1974). In (R. C. Atkinson, D.G. Krantz, R.
C. Luce & P. Suppes. Eds): Contemporary Developments in Mathematical Psychology.
San Francisco: W. H. Freeman.
Pylyshyn, Z. W. (1980). Behav. Brain Sci.
3
, 111-169.
Rajecki, D. W., Lamb, M. E. & Obmascher, P. (1978). Behav. Brain Sci.
1
, 417-464.
Reynolds, V. (1976). The Biology of Human Action.
San Francisco: W. H. Freeman.
Salapatek, P. (1975). In (L. B. Cohen & P. Salapatek, Eds): Infant Perception From Sensation to Cognition,
vol. 1, Basic Visual Processes.
New York: Academic Press, pp. 133-248.
Sarnat, H. B. & Netsky, M. B. (1974). Evolution of the Nervous System.
New York: Oxford University Press.
Saussure, F. de (1959). Course in General Linguistics.
New York: The Philosophical Library.
Schmitt, F. O., Dev. P. & Smith, B. H. (1976). Science
193
, 114-120.
Schwartzman, H. B. (1982). Behav. Brain Sci.
5
, 168-169.
Shaw, R. & McIntyre, M. (1974). In (W. B. Weimer & D. S. Palermo, Eds): Cognition and the Symbolic Process.
Hillsdale, New Jersey: Lawrence Erlbaum, pp. 305-362.
Shepherd, G. M. (1974). The Synaptic Organization of the Brain.
New York: Oxford University Press.
Smith, P. K. (1982). Behav. Brain Sci.
5
, 139-184.
Snyder, S. H. (1980). Science
209
, 976-983.
Spinelli, D. N. (1970). In (K. H. Pribram & D. Broadbent, Eds): The Biology of Memory.
New York: Academic Press, pp. 293-306.
Stent, G. S. (1975). Science
187
, 1052-1057.
Szentagothai, J. (1978). Proc. R. Soc., Ser. B
201
, 219-248.
Tanner, J. M. (1970). In (P. H. Rasmussen, Eds): Carmichael's Manual of Child Psychology.
New York: John Wiley, pp. 77-156.
Thatcher, R. W. & John, E. R. (1977). Foundations of Cognitive Processes.
Hillsdale, New Jersey: Lawrence Erlbaum.
Tribus, M. & McIrvine, E. (1971). In (G. Piel et al., Eds): Energy and Power.
San Francisco: W. H. Freeman.
Ullman, S. (1980). Behav. Brain Sci.
3
, 373-415.
Vanderwolf, C. H. & Robinson, T.E. (1981). Behav. Brain Sci.
4
, 459-514.
Van Valen, L. M. (1982). Behav. Brain Sci.
5
, 294.
von Neumann, J. & Morgenstern, O. (1944). The Theory of Games and Economic Behaviour.
Princeton: Princeton University Press.
Vygotsky, L. S. (1962). Thought and Language.
Cambridge, Massachusetts: MIT Press.
Weiskrantz, L. (1974). In (F. O. Schmitt & F. G. Worden, Eds): The Neurosciences: Third Study Program.
Cambridge, Massachusetts: MIT Press, pp. 189-204.
Werner, G. (1974). In (F. O. Schmitt & F. G. Worden, Eds): The Neurosciences: Third Study Program.
Cambridge, Massachusetts: MIT Press, pp. 171-183.
White, M. J. (1975). Cognitive networks and worldview: the metaphysical terminology of a millenarian community.
Ph. D. dissertation. Department of Linguistics, State University of New York at Buffalo.
Wiener, N. (1961). Cybernetics.
Cambridge, Massachusetts: MIT Press.
Wright, R. V. S. (1978). In (S. L. Washburn & E. R. McCown, Eds): Human Evolution Biosocial Perspectives.
Menlo Park, California: Benjamin Cummings, pp. 215-238.
Yevick, M. L. (1975). Pattern Recognition
7
, 197-213.
Yevick, M. L. (1978). Behav. Brain Sci.
1
, 253-254.